导语
板块一:复杂系统的建立
生物系统的“临界性特点”,在生物进化的过程中,由于基因和基因之间的相互调控以及各种调控元件的介入,使得其保持在一中不稳定且多变的状态。
不同基因的节点与节点之间的相互作用,形成了一套复杂网络化的横向因果关系,在相互作用的集群中,节点输入(基因元件或调控元件)的微小变化可能会引起输出的巨大变化;这些链接之间存在一定的稳态,在变量环境中,由进化和自组织模型介导出现的变异数(基因突变和外源片段插入)。
本文中的RetroMyelin是在最终物种形成后发生的独立入侵事件,随后宿主物种内序列拷贝逐渐多样化;即在稳态状态下引入的变量,在自组织模型和进化调控的模型基础上调控有颌脊椎动物髓鞘的形成;本研究中鉴定到了大鼠RNLTR12-int介导了SOX10调控MBP的形成,进而促进了髓鞘的形成。随后在有颌脊椎动物中鉴定到了RetroMyelin,而且RetroMyelin只能在有颌脊椎动物中检测到,在无颌脊椎动物和无脊椎动物中检测不到。本研究中的模型变量和对应的模型结果如下:
输入变量:RetroMyelin(远古逆转录病毒残余)多次独立入侵
输出结果:趋同进化,促进了髓鞘的形成
模型结果:促进更复杂的大脑结构形成和更多样的物种形态出现
板块二
研究背景
2.1 转座子
概念:一段DNA序列,能够在基因组中通过剪切或转录-逆转录后,在内切酶的作用下,环化后插入另一位点,并对其后的基因起到调控作用。
特征:可以自我复制,可以在染色体上移动;被认为是导致生物基因渐变或突变并最终促使生物进化的根本原因(因此是物种进化的重要痕迹)。转座子的存在说明基因组并不是一个静态的集合,而是一个不断改变自身构成的动态有机体。一部分II型转座子由于“突变积累”失去了转座能力,在进化过程中被“驯化”,参与到基因表达的调控中;转座子在物种基因组中的分布不是随机的,自然选择会筛选对于宿主生存适应的插入片段。
麦克林托克:转座子不仅可以移动,还可以根据特定环境条件或在细胞发育的不同阶段打开或关闭。
分类:根据转座子“跳跃” 方式的不同,分为I型和II型转座子
I型转座子:中间体是RNA,转座子会先被转录为RNA,然后该RNA被逆转录,再次成为DNA,被插入到目标位点中。本研究中的RetroMyelin属于I型转座子。
II型转座子:转座行为是“剪切后粘贴”,转座子两端是两端正向重复序列,接壤的是反向重复序列(回文结构),然后才是中间的插入序列。
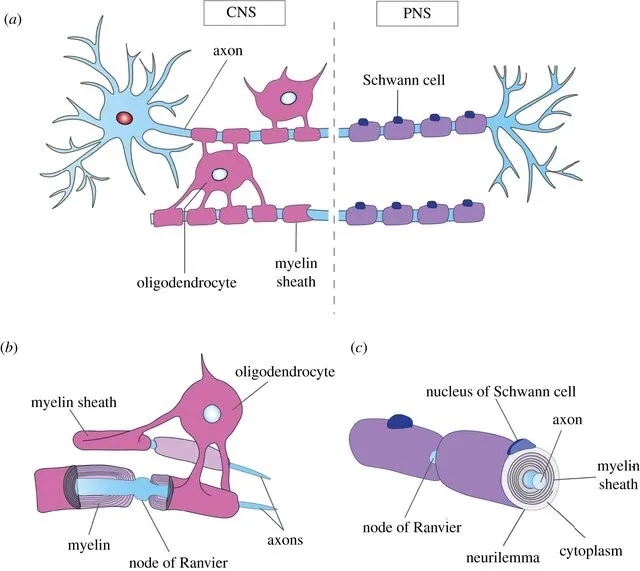
2.2 髓鞘形成
髓鞘是包裹神经轴突的外壳,由中枢神经系统重的少突胶质细胞产生,在有颌动物中首次出现髓鞘,髓鞘的出现使得神经信号可以快速传递,促进更复杂的大脑结构形成和更多样的物种形态出现。MBP:髓磷脂基本蛋白(Myelin Basic Protein,是中枢神经系统(CNS)中的髓鞘的主要成分,本研究中证实了MBP通过与转录因子SOX10结合的RNLTR12-int编码的RNA调控其表达,这一过程对于髓鞘的形成是必要的。
板块三:研究思路
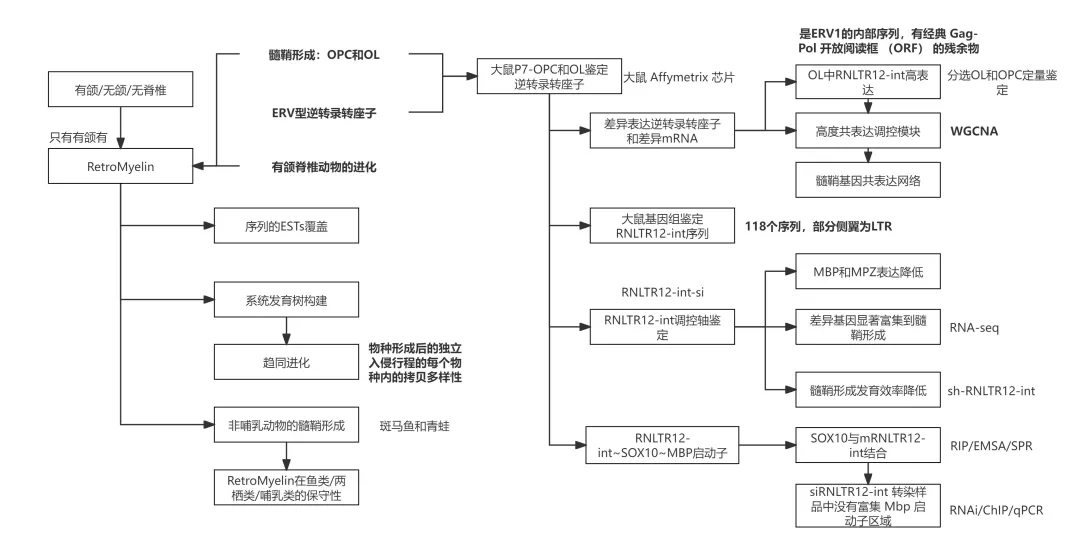
本研究从大鼠大脑分离少突胶质细胞前体(OPC)和少突胶质细胞(OL),借助芯片检测差异逆转录转座子和差异mRNA,通过WGNCA筛选高度相关的调控模块,并借助相对应的免疫沉淀实验,证实RNLTR12-Int~SOX10~MBP的调控路径。并在后续的实验中证实了RNLTR12-Int是一类转座子,在有颌脊椎动物中均存在类似的序列,命名为RetroMyelin;RetroMyelin的功能具有保守性,且只在有颌脊椎动物中存在,对有颌脊椎动物的髓鞘形成和大脑复杂结构的形成至关重要。
板块四:结果概要
4.1 RNLTR12-int是髓磷脂/MBP的潜在调节因子
大鼠p7脑中分离的少突胶质细胞(OPC)和成熟胶质细胞OL进行芯片检测逆转录转座子,鉴定差异逆转录转座子和差异mRNA,与OPC相比,OL中的RNLTR12-int有较高的基因表达水平;为了确定鉴定到的逆转录转座子RNA和蛋白编码mRNA之间的潜在功能关系,进行WGCNA分析。鉴定结果显示有13个高度共调控基因模块,且RNLTR12-int与髓鞘基因共表达网络高度相关。通过调控网络分析,鉴定出RNLTR12-int与MBP存在显著的调控关系。
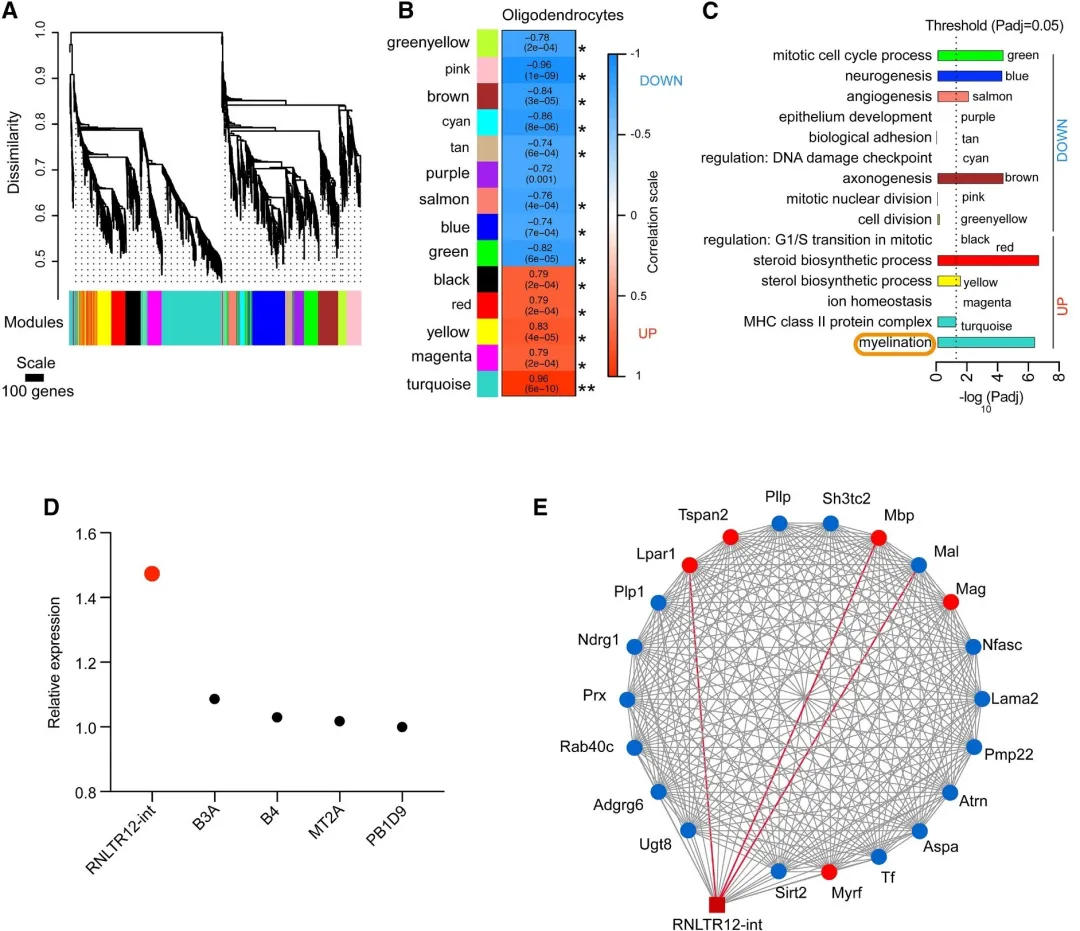
4.2 RNLTR12-int调节发育性髓鞘形成中的Mbp表达
siRNA介导的RNAi,抑制RNLTR12-int的表达,荧光定位结果显示si组的MBP表达会被显著抑制,并限制了OL髓鞘形态的发展。
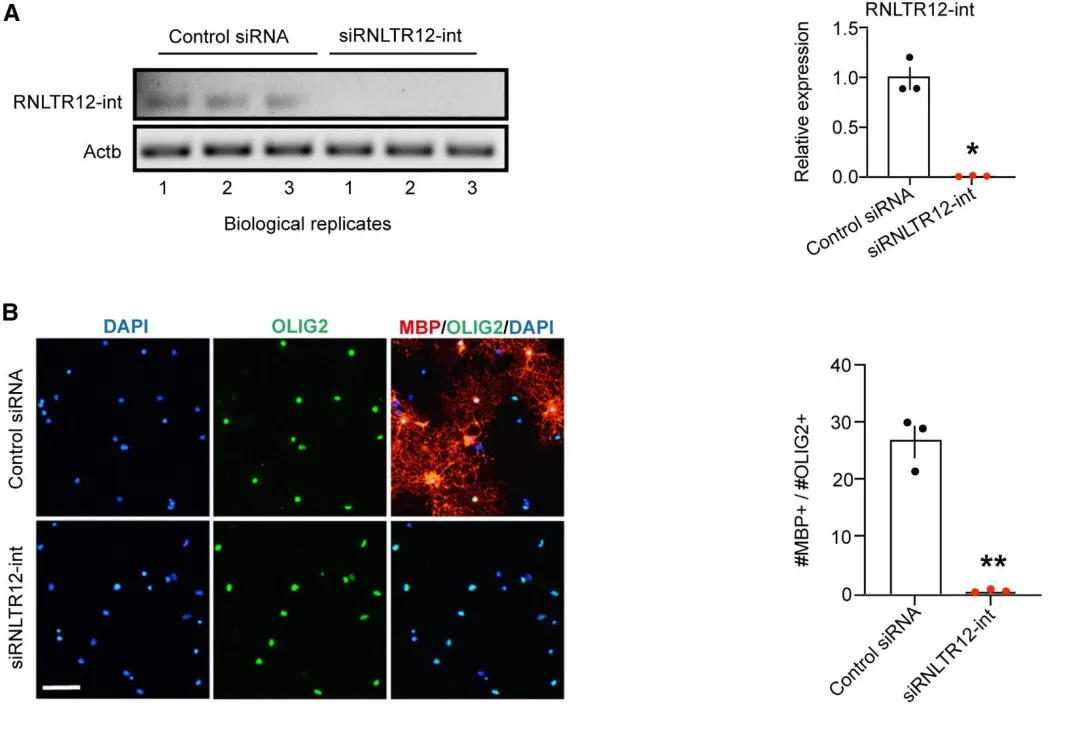
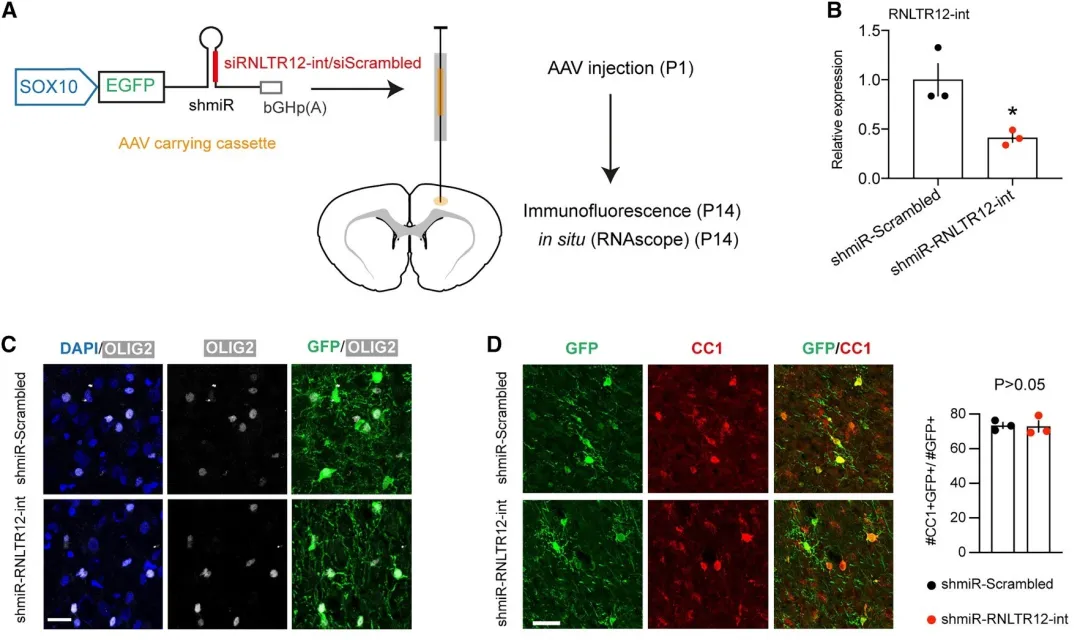
shmiR-RNLTR12-int腺病毒注入大鼠大脑皮层,细胞中OL早期分化标志物CC1和CNP表达未显著降低,即干扰RNLTR12-int并未影响Ols的分化,只影响了髓鞘的形成。转染后的大鼠大脑细胞中的Mbp表达显著降低;电镜结果显示,转染后的OL会产生更多的无髓鞘轴突,表明髓鞘形成效率降低;综上结果表明,MBP表达和体内髓鞘形成对RNLTR12-int的依赖性
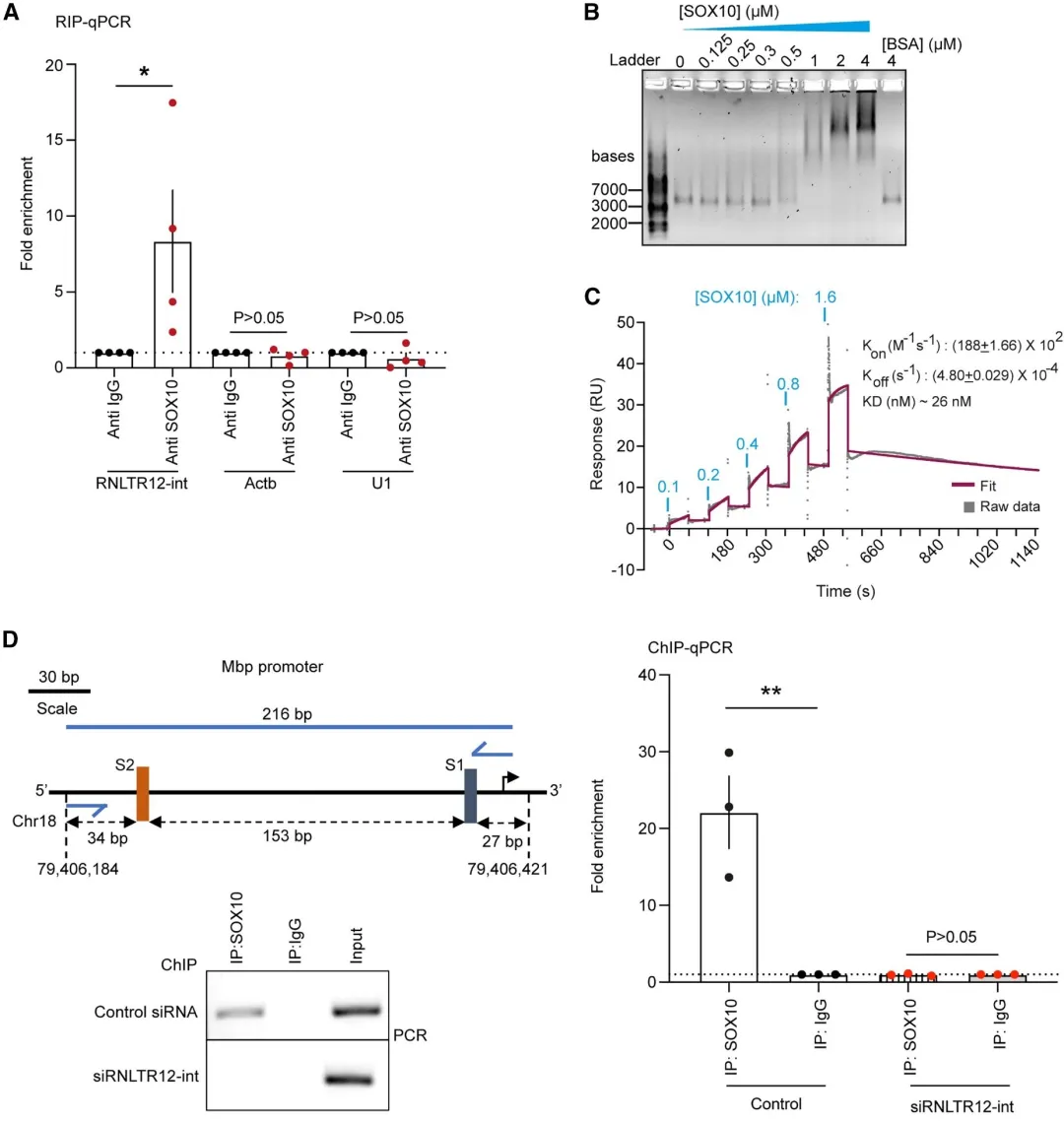
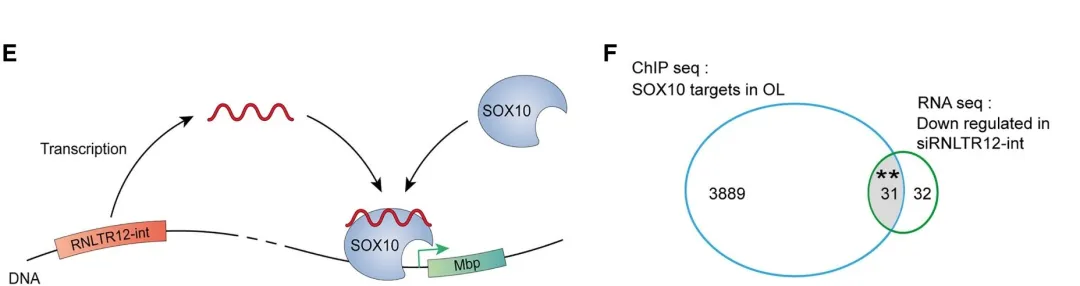
4.3 RNLTR12-int调控SOX10与MBP启动子的结合
大鼠大脑-SOX10抗体进行RIP:结果显示SOX10可以与mRNLTR12-int结合;电泳迁移率位移测定 (EMSA) 和表面等离子体共振 (SPR)证实了SOX1与mRNLTR12-int结合的潜力。需要进一步证实SOX10与Mbp启动子的结合是否依赖于RNLTR12:si RNLTR12-int转染OPC细胞培养,随后进行SOX10-ChIP-qPCR,结果证实SOX10 定位到包括 S2 和 S1 的 Mbp 启动子区域(siRNLTR12-int 转染样品中没有富集 Mbp 启动子区域)。
OL中的其他基因也需要SOX10的调控,比较了RNLTR12-int抑制导致的下调基因列表与OL中潜在的SOX10靶基因;研究发现有31个基因(所有靶基因的0.8%)由于si RNLTR12-int而下调,表明RNLTR12-int可能只调控一部分SOX10的调控(或者说只有0.8%可以主要由RNLTR12-int 调控)。
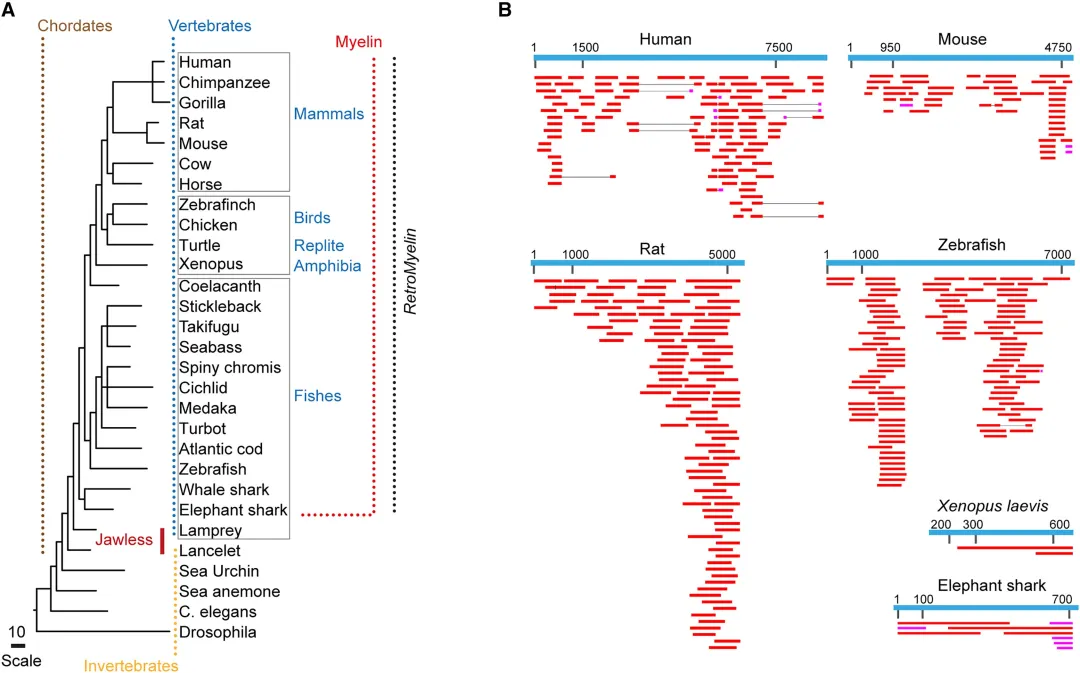
4.4 RetroMyelin只存在于有颌脊椎动物中
基于隐马尔可夫模型在有颌脊椎动物,无颌脊椎动物和无脊椎动物中寻找RNLTR12-int序列,然后进行注释和家族分析。结果显示只能所有有颌脊椎动物中找到类似RNLTR12-int的序列,命名为RetroMyelin(与髓鞘进化相关的逆转录转座子序列)。
从人大小鼠等物种中获取RetroMyelin序列(所有鉴定的RetroMyelin都属于ERV1家族),比对表达序列标签(EST)数据库,发现这些在RNLTR12-int上进行NHMMER搜索后获得的最佳比对结果。
4.5 宿主基因组整合RetroMyelin序列
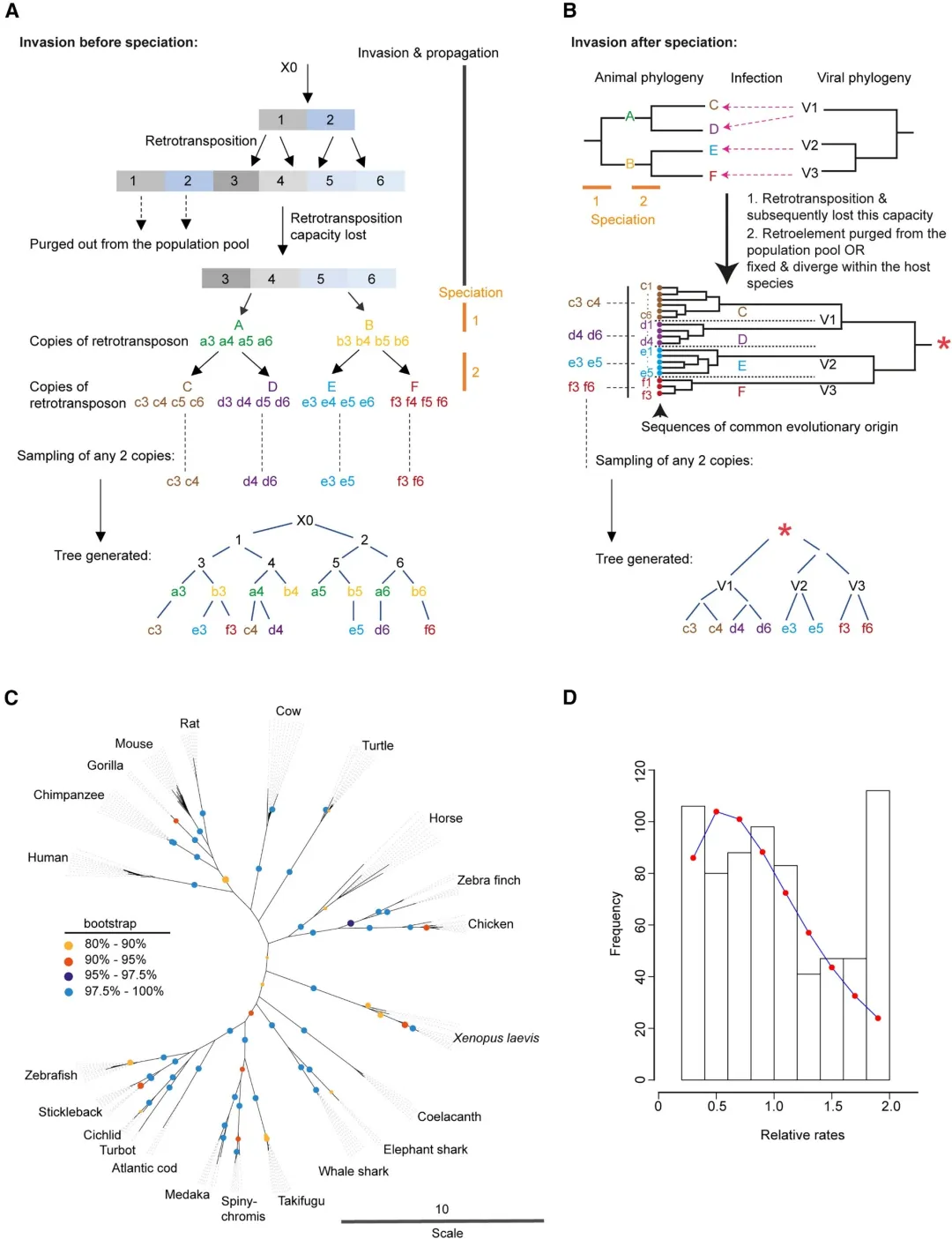
物种进化过程中,RetroMyelin的获得方式一般为两种:
1. 在物种形成之前的共同祖先那里获得:序列分化会从祖先阶段开始,在物种中的拷贝不会聚集到系统发育树上。
2. 在物种形成后的多次逆转录病毒侵入中获得;每个物种的序列拷贝会形成单独的簇(分化只在一个物种内发生)
为了证实RetroMyelin的获得方式,研究选取了22个物种的RetroMyelin拷贝,重构系统发育树,发现每个物种的拷贝都单独聚集在一起,证实了是最终物种形成后发生的单独的入侵事件;每个物种内的序列拷贝的多样性及其对Mbp表达的共同选择可能是通过趋同进化发生的。
研究结果显示进化模型中的进化过程中的选择限制了基本基因组的快速分化,因此文中采用了突变率异质性模型,假设核苷酸替换在位点上的进化效率变化遵循伽玛分布,通过似然比检验(2ẟ = 1,528.5;p < 0.005)证实了突变率异质性的存在。
RetroMyelin的进化相对速率表明(进化速率异质性分析:根据推断的伽玛分布的累积分布函数并绘制计算进化相对速率的期望值),有许多位点(52%)相对保守或正在缓慢进化。
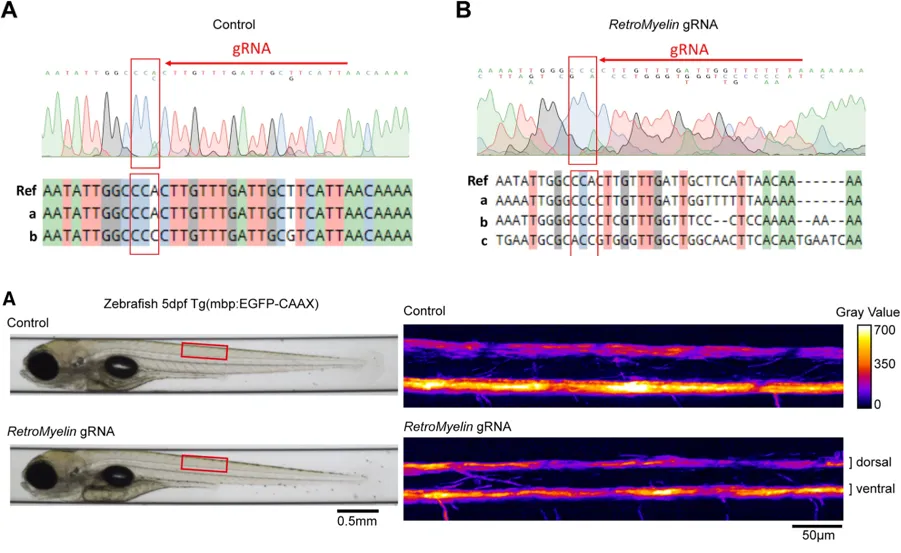
4.6 RetroMyelin调节其他非哺乳类脊椎动物的髓鞘形成
研究为了验证RetroMyelin调节其他非哺乳类脊椎动物的髓鞘形成,分别对斑马鱼和青蛙进行CRISPR-Cas9,编辑破坏RetroMyelin
斑马鱼:与对照组相比,斑马鱼的形态发育明显受到限制,检测到脊髓的Mbp信号减弱(EGFP荧光信号)。
青蛙:MBP信号显著降低。
综上,RetroMyelin在调节鱼类、两栖动物和哺乳动物之间的Mbp转录方面具有保守性。
板块五:结果讨论
5.1 Mbp是髓鞘形成的重要成分
Ols可以形成髓鞘,致密的髓鞘可以加快神经传导,并保持较小的轴突直径,其中MBP是髓鞘形成的重要成分。CNS 和 PNS 的髓鞘包括许多蛋白质,例如髓鞘和淋巴细胞蛋白 (MAL)、MAG、CNP、蛋白脂质蛋白、四跨膜蛋白脂质 (PLP)、TSPAN2、外周髓鞘蛋白 22、脂肪酸结合蛋白 (PMP22) 和 MPZ(一种 Ig 样细胞粘附蛋白)。
上述髓鞘相关蛋白的直系同源物也存在于无颌脊椎动物和无脊椎动物中,并执行其他功能。MBP只存在于有颌脊椎动物中,Mbp的ORF中的部分转录单元也在基因Golli(OL谱系的基因)中存在, Golli存在于七鳃鳗中,但不是MBP的同源物也不是髓鞘形成元素。因此Mbp 的ORF的从头形成是出现致密髓鞘进化出现有颌脊椎动物的重要因素。
5.2RetroMyelin
RetroMyelin存在于所有的有颌脊椎动物中,并在单个物种内表现出高度的序列相似性,并在系统发育树种形成各个物种的不同簇,表明RetroMyelin的获得是物种形成之后的独立入侵。尽管髓鞘形成需要脊椎动物从共同祖先继承许多关键基因,但RetroMyelin在物种形成后的独立获取才是促进髓鞘形成的最核心元素。
RetroMyelin理论上起源于携带RNLTR12-int系列的逆转录病毒的侵入;系统发育分析表明,上述侵入发生在物种形成之后的单独侵入;但是在无颌脊椎动物和无脊椎动物中缺乏固定流程而丢失。在有颌脊椎动物中的多次独立侵入构建了固定的流程,进而形成了拷贝的多样性。
胎盘的合胞素基因来自ERV,通过不同哺乳动物的独立驯化事件,也是趋同进化。ERV1在脊椎动物基因组中的内生化与髓鞘形成的进化出现相结合,这是脊椎动物进化的关键步骤。逆转录病毒感染对于包括人类在内的物种髓鞘进化存在及其重要的影响。
5.3GOF(功能获得实验)
RetroMyelin存在多重拷贝,使得一个基因座损伤,其他拷贝可以充当缓冲剂,确保足够水平的转录表达。髓鞘等复杂性状的进化是一个渐进、复杂的过程,由遗传、环境和选择压力之间的相互作用驱动。需要对RetroMyelin进行遗传固定,并形成新的髓鞘产生网络。
因此通过GOF(功能获得)实验,在七鳃鳗中产生致密髓鞘不仅仅是在短期内引入RetroMyelin和Mbp序列,需要构建一系列的基因表达模块,在七鳃鳗固有的调控流程引入变量(RetroMyelin和Mbp序列),同时还需要综合考虑到自然选择模型和自主模型的总和因素,才能在七鳃鳗中诱导出稳定的髓鞘合成路径。
板块六:参考文献
1.Ghosh, Tanay et al. “A retroviral link to vertebrate myelination through retrotransposon-RNA-mediated control of myelin gene expression.” Cell vol. 187,4 (2024): 814-830.e23. doi:10.1016/j.cell.2024.01.011
2.Lamanna, Francesco, et al. "A lamprey neural cell type atlas illuminates the origins of the vertebrate brain." Nature Ecology & Evolution 7.10 (2023): 1714-1728.
3.Chen, Civia Z et al. “Schwann cell remyelination of the central nervous system: why does it happen and what are the benefits?.” Open biology vol. 11,1 (2021): 200352. doi:10.1098/rsob.200352