编辑 | 水产研究社
最新首发 | 了解最新水产养殖知识,关注:水产研究社
水产养殖,病害防治,水质调节,技术交流,生态养鱼,养虾,养蟹,健康渔业,您值得信赖的水产小帮手。

近些年,由于鱼粉资源的紧缺,水产饲料中植物性原料的用量逐渐增加。
非淀粉多糖(nonstarch polysaccharides.NSPs)和植酸作为植物原料中主要的两大抗营养因子,大量应用后会导致水产动物出现肠道损伤、养分消化率降低等诸多问题。
NSPS主要由纤维素、半纤维素和果胶等多糖构成,不能被机体消化吸收,且会增加肠道中食糜黏度,降低食糜的排空速度,阻碍养分的吸收,并导致肠道中厌氧微生物产生毒素,从而影响动物的健康和生长。
此外,NSPS 作为细胞壁的主要成分,包裹着蛋白质和脂肪等营养物质,但鱼类缺少分解细胞壁的酶,从而干扰鱼类的消化吸收。
植酸是植物原料中磷的主要存在形式,由于鱼类肠道中缺少内源植酸,故对其消化利用率低,大多数的植酸磷被排出体外,会引起水体富营养化。
此外,植酸还能与饲料中的蛋白质和矿物质等结合,降低其消化利用率。
应用酶制剂是提高饲料中营养物质利用的一条有效途径,包括植酸酶、蛋白酶和碳水化合物酶等。
目前,在饲料中补充植酸酶的报道已见于奥尼罗非鱼、黄颡鱼、虹鳟等水产动物中。在罗非鱼和南美自对虾中也有补充碳水化合物酶的报道。
在这些研究中,外源酶制剂的添加抵消了高植物原料所带来的负面影响,提高了水产动物对营养物质的消化利用率,改善了生长性能。
不同酶制剂的作用底物不同,为使酶制剂效果最大化,提高饲料中多种营养物质的消化利用率,常把几种单体酶按一定比例配制成复合酶制剂,增加其协同效应。
以植酸和碳水化合物酶为主的复合酶制剂在肉食性鱼类如欧鳇、鲑鱼和杂食性鱼类如罗非鱼上已有较多研究,而在草食性鱼类上的报道很少。
草鱼作为典型的草食性鱼类,因其生长速度快,抗病性强和肉质鲜美等特点,已成为我国最重要的淡水养殖鱼类之一。
由于草鱼的营养需求较低,故在生产中草鱼饲料的鱼粉用量很低,基本上为全植物性原料。
在这种情况下,如能通过复合酶制剂的使用提高植物性原料的利用率,则可降低生产成本,减少营养物质向环境的排放量。
因此,本试验设计低磷低脂和无鱼粉低磷低脂2种饲料,分别添加以植酸酶和碳水化合物酶为主的复合酶制剂,考察其对草鱼生长、营养物质消化率和沉积率、血清生化指标及肠道组织形态的影响为酶制剂在草鱼饲料中的合理应用提供依据。
1 材料与方法
1.1 试验设计
配制正常鱼粉、磷和粗脂肪含量的常规饲料(正对照)、低磷低脂饲料(负对照1)和无鱼粉低磷低脂饲料(负对照2);
3种饲料中鱼粉、磷酸二氢钙和豆油含量分别为2.0%、1.8%、1.5%;2.0%、1.2%、1.0%和0、1.2%、1.0%。
另在低磷低脂饲料和无鱼粉低磷低脂饲料中分别添加0.6g/kg的复合酶制剂(等量替代次粉),共配成5种试验饲料。
复合酶制剂添加量参考Hlophe-Ginindza等的报道。同时,饲料中添加0.05%的三氧化二钇(Y2O3)作为外源指示剂,用于营养物质消化率的测定。
复合酶制剂主要成分和含量如下:植酸酶1500 000 FYT/kg,木聚糖酶350000FXU/kg,β-葡聚糖酶19250 FBG/kg,果胶酶1750 000 PSU/kg 和淀粉酶800 KNU/kg。
主要饲料原料粗蛋白质含量如下:鱼粉630g/kg,豆粕442g/kg,棉籽粕500g/kg,菜籽粕377g/kg,次粉 169 g/kg,脱脂米糠 143 g/kg。
各主要饲料原料经粉碎过40目筛,充分混匀后,用单螺杆挤压机(SLP-45,中国水产科学研究院渔业机械仪器研究所研制)制成直径2mm的沉性颗粒饲料(制粒温度为90-95 ℃),自然风干后置于-20 ℃冰箱中储存备用。
试验饲料组成及营养水平见表1。

1.2 试验用鱼及饲养管理
取450尾体质健壮的草鱼[平均体重为(16.3+0.2)g],随机分配到15个网箱(1.5 mx1.0 mx1.2 m)中,每个网箱投放30尾。
每3个网箱悬挂于一口室内水泥池(5.0mx3.0 mx1.2 m)中。每日于07:00、12:00和17:00各投喂1次,日投喂量为鱼体重的3%-5%,每天记录饲料消耗量,根据试验鱼的摄食情况适当调整投喂量,以投喂后10分钟内无残饵为宜,各网箱保持基本一致的投喂量。
试验期间养殖水体24h充氧,每5d换水1次,每10d吸污1次,每次换水量约为总水体的1/3。饲养周期60d。
饲养期间水温(29±2)℃,pH7.5-8.0,溶解氧浓度>5mg/L,氨氮浓度<0.2mg/L。试验在上海海洋大学滨海养殖基地进行。
2 结果
2.1 生长性能和形体指标
由表2可知,饲养试验期间,各组均无死亡情况发生,存活率均为100%;
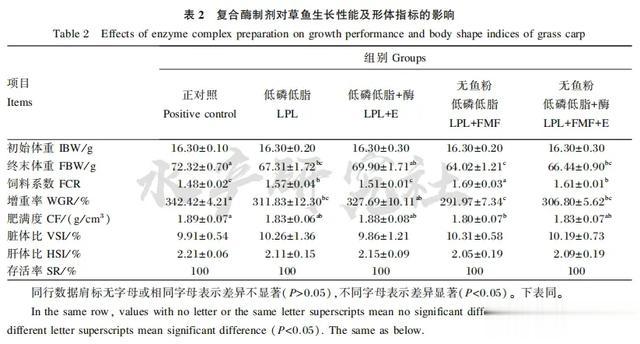
正对照组具有最高的增重率、最高的肥满度和最低的饲料系数,低磷低脂组和无鱼粉低磷低脂组的增重率较正对照组显著降低(P<0.05),而饲料系数则较正对照组显著升高(P<0.05);
在低磷低脂饲料中添加复合酶制剂后,增重率提高15.86%(P>0.10),饲料系数降低0.06(P<0.05),并达到与正对照组基本一致的水平(P>0.05);
在无鱼粉低磷低脂饲料中添加复合酶制剂后,增重率提高14.83%(P>0.05),饲料系数降低0.08(P<0.05),达到与低磷低脂组基本一致的水平(P>0.05);
各组在脏体比和肝体比上无显著差异(P>0.05)。
2.2 体成分和营养物质沉积率
由表3可知,对于各体成分指标,各组在粗蛋白质和粗灰分含量方面无显著差异(P>0.05);

正对照组具有最高的水分,磷含量和最低的粗脂肪含量,低磷低脂组和无鱼粉低磷低脂组的粗脂肪含量较对照组显著升高(P<0.05),而磷含量则较正对照组显著降低(P<0.05);
在低磷低脂饲料和无鱼粉低磷低脂饲料中添加复合酶制剂均显著降低了粗胎肪含量(P<0.05),并使磷含量达到与正对照组基本一致的水平。
由表4可知,低磷低脂组和无鱼粉低磷低脂组的蛋白质和灰分沉积率较正对照组显著降低(P<0.05)而脂肪沉积率则较正对照组显著升高(P<0.05);

在低磷低脂饲料和无鱼粉低磷低脂饲料中添加复合酶制剂后,蛋白质和磷沉积率显著增加(P<0.05),脂肪沉积率显著降低(P<0.05)其中低磷低脂+酶组的磷沉积率甚至显著高于正对照组(P<0.05)。
2.3 肠道消化酶活性和营养物质消化率
由表5可知,低磷低脂组和无鱼粉低磷低脂组的肠道蛋白酶活性以及干物质、蛋白质和磷消化率均显著低于正对照组(P<0.05);

在低磷低脂饲料和无鱼粉低磷低脂饲料中添加复合酶制剂后,肠道蛋白酶活性以及干物质、蛋白质和磷消化率均显著增加(P<0.05),肠道淀粉酶活性在数值上也有所增加(P>0.05)。
2.4 血清生化指标
由表6可知,与正对照组相比,低磷低脂组血清碱性磷酸酶活性显著增加(P<0.05)无鱼粉低磷低脂组的血清碱性磷酸酶、谷草转氨酶和谷丙转氨酶活性均显著增加(P<0.05);

在低磷低脂饲料和无鱼粉低磷低脂饲料中添加复合酶制剂后,血清碱性磷酸酶和谷丙转氨酶活性显著降低(P<0.05);各组间血清磷含量无显著差异(P>0.05)。
2.5 肠道组织形态
由表7可知,无鱼粉低磷低脂组的肠道绒毛高度和绒毛纵截面面积均较正对照组显著降低(P<0.05),但在无鱼粉低磷低脂饲料中添加复合酶制剂后,肠道绒毛高度和绒毛纵截面面积均显著升高(P<0.05),达到与正对照组基本一致的水平;

各组的肠道肌层厚度无显著差异(P>0.05)。正对照组、无鱼粉低磷低脂组和无鱼粉低磷低脂+酶组的肠道组织切片见图1。
3 讨 论
3.1 复合酶制剂对草鱼生长性能和营养物质利用的影响
Hlophe-Ginindza等在罗非鱼饲料中添加0.5 g/kg以植酸酶和碳水化合物酶为主的酶制剂后显著提高了蛋白质消化率、肠道蛋白酶活性以及生长性能。
Kim 等和 Selle 等在饲料中添加以植酸酶和碳水化合物酶为主的酶制剂分别显著提高了猪和鸡的蛋白质消化率和生长性能。
本试验中,在低磷低脂饲料和无鱼粉低磷低脂饲料中添加复合酶制剂后,显著提高了干物质、蛋白质和磷消化率,有提高生长性能的趋势。
本试验所用复合酶制剂由植酸酶、木聚糖酶、β-葡聚糖酶、果胶酶和淀粉酶组成,其作用底物分别为植酸、本聚糖、β-葡聚糖、果胶质和淀粉,不同的酶制剂作用机制不同。
通常认为,酶制剂可以通过改变肠道微生物组成影响营养物质的消化吸收,最终改善生长性能。
植酸酶可降解植酸,提高植酸盐的利用率,增加肠道中微生物繁殖所必需的磷,进而调节肠道微生物的组成。
Adeoye等在饲料中添加以植酸酶为主的酶制剂饲喂罗非鱼发现肠道中邻单胞菌属数量增加,面鲸杆菌属数量减少。
此外,植酸酶还可以增加猪和鸡肠道中双歧杆菌属和乳酸杆菌属等有益菌的数量。
木聚糖酶、β-葡聚糖酶和果胶酶等碳水化合物酶,一方面可以分解细胞壁,降低肠道食糜黏度;
另一方面可以降解生成功能性寡糖抑制肠道中病原菌的定植,增加益生菌的数量。
Jiang 等研究发现,碳水化合物酶增加了建鲤肠道乳酸杆菌属的数量,降低大肠杆菌和气单胞菌属的数量。
饲料中添加碳水化合物还可以提高草鱼肠道芽孢杆菌属的数量,降低链球菌属和假单胞菌属等的数量。
本试验所用复合酶制剂由植酸酶和多种碳水化合物酶组成,它们各自作用于不同底物,理论上应优于单一添加的效果。
然而,也有报道认为酶制剂的联合使用较单独使用并没有产生额外的益处,甚至会降低营养物质利用率;Meng等认为酶制剂的联合使用效果可能与饲料组成有直接关系。
由于本次试验未设置单独添加植酸酶和碳水化合酶的组,故难以分辩出这2种酶各自的贡献度,今后将进一步研究单体酶之间的相互作用。
3.2 复合酶制剂对草鱼体成分的影响
本试验中低磷低脂组和无鱼粉低磷低脂组的全鱼粗脂肪含量均显著高于正对照组,这是因为磷的缺乏可增加组织内脂质的沉积。
在磷缺乏情况下,游离脂肪酸酯化形成脂酰辅酶 A 会被抑制,降低脂肪供能,同时肝脏中谷丙转氨酶活性和糖异生作用增强,使脂肪利用率降低,最终导致鱼体粗脂肪含量增加。
饲料中添加复合酶制剂后,在其中的植酸酶的作用下,磷消化率和沉积率得到显著提高,满足了鱼体对磷的需求,脂肪代谢恢复正常,使得鱼体粗脂肪含量下降。
植酸酶可以降解植酸、提高磷利用率的报道也见于虹鳟,狼鲈和斑点叉尾鮰等鱼类。
此外,本试验在低磷低脂饲料和无鱼粉低磷低脂饲料中补充复合酶制剂后,对全鱼粗蛋白质含量没有产生显著影响,但在一定程度上改善了生长性能,显著提高了蛋白质沉积率。
Maas等和 Ai 等在饲料中添加植酸酶和碳水化合物酶的复合酶制剂后,同样也提高了鱼体氮沉积率和生长性能。
这可能是因为酶制剂的添加使营养物质消化率得到提高,生长性能得到改善,蛋白质效率提高,使得蛋白质沉积率增加,由于各组饲料营养水平差异并不大,加上鱼类的自我调节能力,使得各组鱼体粗蛋白质含量并没有出现显著差异。
3.3 复合酶制剂对草鱼血清生化指标的影响
通常情况下,血清谷丙转氨酶,谷草转氨酶和碱性磷酸酶可作为评估肝脏状况的指示酶。
碱性磷酸酶由成骨细胞分泌,反映了成骨细胞的活跃程度和机体的磷营养状况。
本试验中,没有发现无鱼粉低磷低脂组草鱼出现花肝、绿肝或鱼体畸形等症状,说明磷缺乏的程度并不严重,但血清碱性磷酸酶活性升高,表明机体可能动员了体内的磷贮备(骨磷),以保证其他组织对酶的需求。
通常,血清碱性磷酸酶活性更易受饲料磷水平的影响,当饲料中磷缺乏时血清碱性磷酸酶活性升高,这在郑涛等对奥尼罗非鱼和张晓清等对建鲤的研究中已有报道;
当添加酶制剂(植酸酶)后,磷的利用率得到提高,缓解了磷缺乏的状况,故碱性磷酸酶活性恢复正常。
谷丙转氨酶,谷草转氨酶是机体分布最广,活性最强的转氨酶。
和哺乳动物一样,鱼类的组织或器官受到功能性损伤时,某些特有的细胞会释放到血液里面被检测出来。
谷草转氨酶主要存在于肝脏,谷丙转氨酶在肝细胞受损时会释放到血液中,血清中谷丙转氨酶活性的相对增加能反映肝功能障碍。
本试验中,无鱼粉低磷低脂组草鱼的血清谷草转氨酶和谷丙转氨酶活性均较正对照组显著升高,饲料中添加复合酶制剂后,血清中谷丙转氨酶活性显著降低,表明草鱼的肝脏功能有所恢复,但血清中谷丙转氨活性的降低不显著,这可能是谷丙转氨酶比谷草转氨酶更敏感的缘故。
3.4 复合酶制剂对草鱼肠道组织形态的影响
肠道组织形态与其消化吸收功能密切相关。
通常,肠绒毛高度和截面积的增加可以增大与食物的接触面积,从而促进消化,肌层厚度则可反映肠道的收缩能力。
本试验中,低磷低脂组的绒毛高度和绒毛纵截面面积较正对照组在数值上有所降低,而无鱼粉低磷低脂组的绒毛高度和绒毛纵截面面积则显著低于正对照组;
一方面可能是饲料中酶不足和缺少鱼粉来源的某些未知因子而产生的后果,另一方面可能是植物原料中较多的NSPS 使得食糜在肠道中停留时间过长,导致有害微生物大量繁殖损害肠道。
在无鱼粉低磷低脂饲料中添加复合酶制剂后,上述问题得到缓解,故肠道绒毛高度和绒毛纵截面面积均显著增加。
Ramos 等在低鱼粉饲料中添加以植酸和碳水化合物酶为主的酶制剂,显著降低了鲻鱼肠道病变的几率,改善了肠道健康。
研究发现,在饲料中添加以植酸酶和碳水化合物酶为主的酶制剂后显著提高了罗非鱼中肠绒毛密度,而单独添加植酸酶或碳水化合物酶后均显著提高了罗非鱼中肠绒毛密度和黄颡鱼前肠绒毛高度。
此外,在猪和鸡的试验中,饲料中添加碳水化合物酶和植酸酶也显著改善了肠道组织结构。
本试验中无鱼粉低磷低脂+酶组的肠组织结构的改善与碳水化合物酶和植酸酶均有关系,具体各自的贡献度有待于今后进一步研究。
结 论
本试验条件下,在低磷低脂饲料和无鱼粉低磷低脂饲料中补充0.6g/kg 的复合酶制剂均可提高草鱼的蛋白质和磷沉积率以及干物质、蛋白质和磷消化率,降低饲料系数,从面提高营养物质利用率,改善生长性能;
此外,在无鱼粉低磷低脂饲料中补充0.6g/kg的复合酶制剂还可增加草鱼肠绒毛高度和截面积,从面改善肠道组织形态。
