远端关节挛缩症的特征是手脚远端区域的挛缩。严重的类型是远端关节弯曲2A 型(也称为 Freeman-Sheldon 综合征)和 2B 型(也称为 Sheldon-Hall 综合征),会出现面部受累和脊柱侧凸,而 AMCD1 (DA1) 则没有。AMCD1又叫做1 型远端关节挛缩,影响大约万分之一的儿童,是最常见的类型型。
肌节中编码蛋白质的多个基因,包括肌球蛋白重链 3 (MYH3)、肌球蛋白重链 8 (MYH8)、原肌球蛋白 2 (TPM2)、肌钙蛋白 I2 (TNNI2)、肌钙蛋白 T3 (TNNT3) 和肌球蛋白结合蛋白 C1 (MYBPC1) ),与远端关节弯曲综合征有关。尽管 MYH3 突变导致几乎所有 Freeman-Sheldon 综合征(2A 型)病例和近三分之一的 Sheldon-Hall 综合征(2B 型)病例,但根据佳学基因检测的记录,MYH3 突变尚未在远端关节弯曲类型1的患者中发现。先前的基因检测记录表明,已知的 1 型远端关节挛缩症的基因突变很少,因此,基因解码认为这一疾病存在遗传异质性,并且仍有待确定其他致病基因。
由于涉及远端关节弯曲综合征的基因数量众多且相对较大,因此需要对所有基因进行测序的新方法,以有效地确定个体家族中远端关节弯曲的遗传基础。外显子组测序是一种新的基因测序工具,它使用下一代测序方法来识别整个人类基因组(“外显子组”)编码外显子内的突变。由于外显子组仅占基因组的 1%,因此目前仅对外显子进行测序的成本只是全基因组测序成本的一小部分。
致病基因鉴定基因解码采用的是基于全外显子测序的结构功能分析法。其中的全外显子组测序使用的是下一代外显子组捕获测序方法,代表了致病基因鉴定方法的重大进步,尽管存在局限性,但这些方法可能很快会对遗传性肌肉骨骼疾病患者的诊断产生重大影响。例如,在本例中描述的家族中对 MYH3 突变的鉴定扩大了与 MYH3 突变相关的表型,包括 AMCD1,并表明 1、2A 和 2B 型之间可能存在大量重叠。孤立性马蹄内翻足的鉴别诊断应考虑 AMCD1,特别是在患者或家庭成员存在轻微手部挛缩的情况下。
采用外显子测序基因检测明确关节弯曲是一种AMCD1遗传病材料和方法DNA 是从一个家族的所有受影响和几个未受影响的成员利用抗凝EDTA采血管采集的静脉血。该家族包括 6 名 1 型远端关节挛缩症患者。使用 SureSelect All Exon 38Mb 试剂盒(Agilent Technologies)对其中一名受影响的家庭成员(先证者)进行外显子组捕获)。在 HiSeq 2000 测序仪(Illumina,San Diego,California)的单通道上对 DNA 进行测序。使用 Novoalign 软件(Novocraft Technologies,Selangor,Malaysia)将序列与基因组的 hg18 组装比对,并使用 SAMtools 和 SeattleSeq识别突变的基因序列。在分析时,仅包含至少六倍读取覆盖率和 Phred 标度单核苷酸多态性质量≥30 的变体用于进一步分析。存在于 1000 基因组计划 ( http://www.1000genomes.org )、dbSNP(build 129)或20名无关疾病患者的内部外显子组数据库中的变异已被删除. 使用 ABI 3730 测序仪(Life Technologies,Carlsbad,California)通过 Sanger 测序对所选的突变进行验证。
假设 80% 的外显率和 0.1% 的频率导致疾病的等位基因,使用来自六个受影响和六个未受影响的家庭成员的阵列基因分型数据进行连锁分析。基因分型佳学基因使用使用 Mapping 10K XbaI 阵列进行。然后进行进行连锁分析。
采用外显子测序基因检测明确关节弯曲是一种AMCD1遗传病结果一个多代家庭出现不完全外显性、常染色体显性遗传的 AMCD1,其特征是马蹄内翻足和轻度手部挛缩。5 位家庭成员患有马蹄内翻足,其中 4 人为双侧,1 人为单侧(图 1A)。患有单侧马蹄内翻足的先证者(家谱中鉴定为 III:3)采用 Ponseti 方法(图 1B) 但有两次复发,需要重复跟腱切断术以保持矫正。其他患有马蹄内翻足的家庭成员接受了广泛的软组织松解治疗。两名患有马蹄内翻足的家庭成员(个体 II:7 和 II:9)也有轻度手部挛缩,主要累及远端指间关节(图 1C和D)。此外,一名家庭成员(个人 II:1)在青春期出现远端指间关节挛缩,但没有马蹄内翻足或其他下肢受累。先证者和其他仅表现出马蹄内翻足的个体没有手部受累的其他迹象,包括没有屈曲折痕、压倒手指和拇指近端。由于没有面部受累,因此诊断为 AMCD1。

图1:1型远端关节挛缩症家族成员的远端肢体挛缩。A:先证者的脚(鉴定为III:3 in图 2), 显示左脚单侧马蹄内翻足。B:使用 Ponseti 方法矫正马蹄内翻足后先证者的脚,在 7 岁时显示出有限的残余畸形。C:先证者阿姨的近端拇指和远端指间关节挛缩(确定为 II:7 in图 2). D:先证者父亲的弯曲指畸形(鉴定为 II:9 in图 2).
肌球蛋白结合蛋白 C1 (MYBPC1) 的测序是一种最近与 1 型远端关节挛缩症的某些病例有关的基因,未发现任何致病突变。然而,由于几乎没有发现导致 AMCD1的基因,AMCD1致病基因鉴定基因解码在对先证者的 DNA 进行了基于外显子组测序基因检测的基因解码过程。
对 Illumina HiSeq 配对末端序列的单个通道的分析产生了 142,366,935 个读数,其中 93% 可以与人类基因组对齐。这在 6× 的最小测序深度下提供了 93% 的序列覆盖率。使用这种方法,在先证者的外显子组中识别出总共 24,323 个会导致非同义氨基酸取代、剪接位点变异、插入或缺失的变异。在排除对照数据库中出现频率 >5% 的变体后,仍然存在 524 个单核苷酸基因序列突变和 49 个插入缺失(插入或缺失)。
这 573 个基因突变序列中的一个由 MYH3 中的错义突变组成,MYH3 是一种胚胎肌球蛋白重链基因,之前曾与 2A 型和 2B 型远端关节弯曲有关。Sanger 测序证实了所有患有马蹄内翻足的个体以及患有青春期手部挛缩症的个体都存在 MYH3 F437I 突变(图 2). chr17:10488494 (hg18) 处的 A→T 错义突变导致氨基酸异亮氨酸取代残基 437 处的苯丙氨酸。该残基位于肌球蛋白头部内,在多个物种中高度保守 (图 3),但它位于 ATP 结合槽之外,突变似乎会导致最严重的远端关节弯曲,即 Freeman-Sheldon 综合征(2A 型)(图 4) 。PolyPhen-2 程序以 99.7% 的置信度预测这种突变“可能具有破坏性”。

图 2:该家族的系谱,显示有和没有 AMCD1的成员。黑色阴影表示马蹄内翻足,星号表示手部挛缩。MYH3 F437I 突变的存在用“Mut”表示,不存在用“WT”表示。

图 3:多个物种序列中与 F437I 突变对应的位置周围的 MYH3 氨基酸比对。残基 437 处的苯丙氨酸由矩形表示,并且在所示的所有物种中都是保守的,包括小鼠、大鼠、人、牛、青蛙和鸡。使用 CLUSTALW 程序 ( http://align.genome.jp ) 对序列进行比对。

图 4:包含氨基酸 4 到 1022 的 MYH3 S2(肌球蛋白头)区域的带状图。在具有 AMCD1的家族中鉴定的 MYH3 F437I 突变的位置以橙色显示。导致 2A 型远端关节弯曲(Freeman-Sheldon 综合征)的其他五个已知突变的位置以红色显示;其中四个位于肌球蛋白头部并位于形成 ATP 结合位点凹槽中。该图像由 Swiss-PDBViewer (DeepView) 程序 ( http://ca.expasy.org/spdbv ) 为蛋白质登录号P11055 (MYH3)生成。
为了确认此 MYH3 F437I 突变的重要性,使用来自该家族所有六个受影响个体的 Affymetrix 10K 单核苷酸多态性数据进行了连锁分析。最大的 LOD(对数比值)得分为 1.86,位于包含 MYH3 基因的 17p 号染色体上(图 5), 而 MYH3 突变是唯一位于大约 3 Mb 连锁区间内的外显子组变异。其他不太重要的连锁区间均不包含任何其他罕见变体。
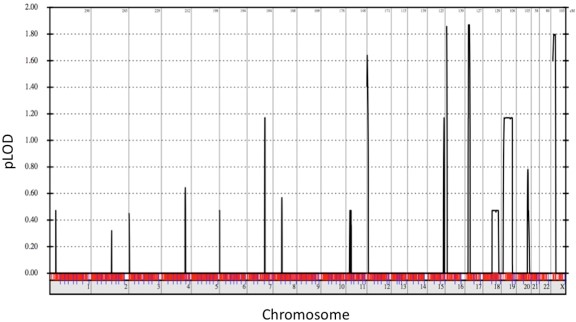
图 5:1 型远端关节挛缩症与染色体 17p 的联系。从 Affymetrix 10K 阵列数据构建的 1 型远端关节挛缩症家族的参数化多点 LOD(几率的对数)分数的全基因组概况。从 p 端(左)到 q 端(右)的人类染色体在水平轴上连接在一起。垂直轴显示了水平轴上指示的人类染色体上物理位置的参数 LOD 分数。1.86 的峰值 LOD (pLOD) 得分(定义最强的连锁区域)位于染色体 17p 内。
采用外显子测序基因检测明确关节弯曲是一种AMCD1遗传病材料和方法讨论我们的结果表明,MYH3 突变可能与 1 型远端关节挛缩症(以及如前所述的 2A 和 2B 型4)相关。所有类型的远端关节弯曲都以家族之间和家族内广泛的表型变异为特征1,我们的数据表明,远端关节弯曲 1 型与 2A 和 2B 型也可能存在临床和遗传重叠。需要考虑基因型和表型的这些综合征的未来分类系统。
将 MYH3 鉴定为该 AMCD1家族中致病突变的位置得到了该基因与 2A 型和 2B 型远端关节弯曲的已知关联的支持,这些疾病与 AMCD1密切相关,并且通过预测 F437I 突变会破坏蛋白质的功能。此外,残基 437 处的苯丙氨酸在许多哺乳动物物种中高度保守,并且未在任何对照中发现突变。最值得注意的是,所有受影响的家庭成员中 MYH3 F437I 突变的分离增加了我们对正确识别致病突变的信心。对其他家族成员的变异测序仍然是有用的,如果不是关键的话,
正如 Toydemir 等人所建议的,MYH3 突变的位置可能与远端关节弯曲的严重程度相关。 在所有受影响的家庭成员中,远端关节弯曲的持续轻微表型进一步支持了涉及 MYH3 的疾病的基因型-表型相关性。F437I 突变略微位于 ATP 结合沟外,导致最严重形式的远端关节弯曲、Freeman-Sheldon 综合征(2A 型)的突变位于. 然而,由于缺乏方便的检测方法,了解可能改变骨骼肌收缩力(通过影响核苷酸结合、ATP 催化或与肌动蛋白或调节蛋白的相互作用)的 MYH3 错义突变的生理影响一直受到阻碍。需要进一步研究以更好地了解这些突变如何导致疾病。
本研究中描述的家庭还表明,将 AMCD1确定为孤立性马蹄内翻足的原因偶尔会遇到困难。尽管该家族中有 3 人患有孤立性马蹄内翻足,但即使有 1 名家族成员存在手部挛缩,也应表明存在远端关节弯曲的可能性。然而,手部挛缩可能要到青春期才会出现,正如在具有 MYH3 F437I 突变的个体 II:1 中所证明的那样。虽然如果不使用适当的治疗(例如支具)可能会导致挛缩恶化,但通常认为挛缩在远端关节弯曲的病例中是非进展性的. 远端关节挛缩症中挛缩的非进行性且通常是先天性的性质归因于基因缺陷的位置,例如 MYH3,据信这些基因仅在胎儿发育期间表达。然而,最近的研究表明,MYH3 也在成人肌肉中表达,也许可以解释这个家族中一名成员的迟发性手部挛缩。在高度外显性、常染色体显性马蹄内翻足家族的鉴别诊断中也应考虑远端关节弯曲,因为这种遗传模式在孤立的马蹄内翻足中相对罕见,除了与复发性染色体 17q23 微缺失和微复制相关的马蹄内翻足外,大约导致所有家族性马蹄内翻足病例的 5%。对治疗有抵抗力的马蹄内翻足也应该提示远端关节弯曲的可能性。尽管先证者的马蹄内翻足使用 Ponseti 方法进行了非手术治疗,她出现了多次复发。尽管存在这些并发症,但使用 Ponseti 方法治疗远端关节弯曲的患者仍有可能获得良好的结果。
这项研究的结果证明了新的测序方法在诊断人类肌肉骨骼疾病方面的能力。虽然本研究中患者的外显子组测序是在研究基础上进行的,但临床可用的诊断测试将很快利用类似的方法来筛选大基因组以寻找与疾病相关的突变。出于多种原因,外显子组分析是识别致病突变的合理方法。首先,目前发现的绝大多数致病突变都位于基因的编码区(外显子组)内,尽管这种情况在未来可能会发生变化,因为研究人员现在正在发现不编码蛋白质的区域的范围会影响人类健康和疾病. 其次,由于外显子组仅占基因组的 1%,因此目前对外显子组测序的成本只是全基因组测序成本的一小部分。最后,外显子组分析不如整个基因组分析复杂,因为可以使用成熟的方法来预测或通过实验测试突变对蛋白质功能的影响。
外显子编码变异的解释虽然比全基因组序列分析结果的解释简单,但仍然很复杂。外显子组数据的分析在很大程度上依赖于使用过滤方法来排除一些健康(对照)个体中也存在的变异。这些正常变异是通过 1000 人基因组计划的努力编制的,该计划的目标是在来自不同人群的 1000 多名健康个体中识别罕见变异. 然而,此类“对照”数据库可能包括具有低外显率的致病突变以及导致迟发性疾病的突变,这些突变尚未在提供对照样本的据报道正常个体中表现出来。人类基因突变数据库等数据库中的致病变异集合也将有助于分析外显子组测序数据,但该数据库目前仅列出了 13 个 MYH3 突变. 由于导致远端关节弯曲等罕见疾病的突变可能仅在单个家族中发现,因此此类数据库目前在外显子组分析中的用途可能有限。然而,最终,它们可能会充满已知与特定疾病相关的高度可信的变体。最后,算法可用于预测修饰蛋白质的基因突变的功能影响。然而,许多与疾病相关的突变,包括先前描述的人类肌球蛋白重链基因、中的几个突变,被 PolyPhen-2 等程序错误地预测为“良性”突变,从而限制了此类分析的整体用途.
尽管外显子组测序存在局限性,例如难以将致病变异与已识别的大量其他新变异区分开来,但我们预计这些技术将迅速推进医学研究并彻底改变患者护理。正如此处 MYH3 所证明的那样,随着外显子组测序技术识别在相关疾病患者中也发现的突变,疾病相关基因变异逐渐被证明是导致表型范围扩大的原因。尽管 MYH3 突变是导致孤立性马蹄内翻足的罕见原因,当任何家庭成员出现手部挛缩(即使是青春期出现的挛缩)或马蹄内翻足严重且难以治疗时,应考虑远端关节弯曲的诊断。尽管 MYH3 测序目前可作为单基因基因检测,但我们预计扩展基因组或基于外显子组的测序将很快可用于肌肉骨骼疾病,如远端关节弯曲。
