文:回溯档案
编辑:回溯档案
棉花是最重要的经济作物之一,在全球纺织品市场上供应的天然纤维数量最多,棉属(棉属)由近50种组成,其中2种,包括2种二倍体(G. arboreum和G. herbaceum,26n = 2× = 4 AA)和52种异体四倍体(G. hirsutum和G. barbadense,1n = 2× = 3 AADD),已被广泛栽培,具有纤维的商业价值。

陆地棉(G. hirsutum)是棉花品种中种植面积最大的,提供了现代纺织工业所需的大部分有价值的纤维,纤维发育分为四个重叠阶段,即纤维起始、生长、次生细胞壁增厚和成熟。
棉纤维被认为是一种强大的细胞研究模型,因为它是一种易于分离的单细胞,具有不同的棉纤维细胞发育阶段,可以进行体外胚珠培养,以进一步研究棉纤维的详细发育阶段。

碱性螺旋环螺旋/螺旋环螺旋(bHLH/HLH)蛋白以碱性螺旋环螺旋结构域命名,bHLH结构域由50-60个氨基酸组成,具有两个功能不同的区域,N末端碱性区域和C末端螺旋-环-螺旋区域。
碱性区域由大约15个氨基酸和许多碱性残基组成,可以识别并与DNA结合,HLH区,包括两个两亲性α螺旋,由具有可改变序列的环区分开,可以与其他HLH结构域形成二聚体。

在高等植物中,bHLH/HLH蛋白参与植物生长发育的调节,如光信号传递、植物激素信号和器官发育,例如在光信号传输中,AtPIF4主要通过与光致色素相互作用来调节光形态发生。

AtSPT不仅与拟南芥不育有关,而且与DELLA蛋白具有协调关系,共同调节赤霉素(GA)反应基因的表达,AtSPT突变可以消除其对细胞生长的抑制AtGL3还参与拟南芥毛状体的发展,通过形成促进comlpex TTG1 / GL3(EGL3)/GL1的毛状体。
在拟南芥的茎或叶表皮细胞中,复合物可以诱导下游基因GL2的表达,以确定表皮细胞的发育,而MYB蛋白AtCPC、AtTRY、AtTCL1和AtETC1可以与GL3结合,与GL1竞争,从而负调节毛状体的起始。

油菜素类固醇(BR)信号传导是拟南芥中一种描述良好的信号通路,在植物生长发育中起着至关重要的作用,许多bHLH/HLH蛋白参与BR信号转导。
它们中的大多数可以形成同源二聚体和异二聚体,赋予它们在调节多个转录程序中起作用的能力,例如AtCESTA可与AtbHLH044(BEE1)形成异二聚体,正调控BR生物合成相关基因CPD的表达。

AtbHLH064(HBI1)通过调节下游基因表达促进细胞生长,其功能可被AtbHLH158(IBH1)抑制,AtbHLH136(PRE1)和AtbHLH158(IBH1)的相互作用可缓解抑制。
一份报告还表明,AtbHLH136(PRE1),AtbHLH158(IBH1)和AtbHLH049/074/077(ACE1 / 2 / 3)构成了竞争性调节细胞生长的三角拮抗bHLH系统。

外源性应用BL(BR)促进棉纤维细胞伸长,而用Brz(一种BR抑制剂)处理棉花芽导致完全没有纤维细胞分化,表明纤维起始和伸长需要BR。
此外,我们之前的研究表明,Gh14-3-3蛋白通过与GhBZR1相互作用来调节BR信号传导,从而参与调节纤维的起始和伸长。

此外,棉花bHLH/HLH转录因子可能在纤维发育中发挥重要作用,然而到目前为止,人们对bHLH/HLH转录因子如何在棉花纤维发育过程中调节BR信号传导知之甚少。
在我们的研究中,我们从基因组上广泛鉴定了可能参与纤维发育中BR信号传导的棉花bHLH / HLH基因,此外还详细研究了这些棉花bHLH/HLH转录因子的特征。
棉花bHLH/HLH转录因子的表征
为了鉴定旱地棉花(G. hirsutum)基因组中的bHLH/HLH转录因子基因,所有已发表的拟南芥和水稻bHLH/HLH基因序列都用作查询,对棉花基因组数据库。
最初在棉花中鉴定出498个候选bHLH / HLH基因,其中丢弃了61个重复序列,此外为了评估初始结果的可靠性,根据报告的方法,使用Hmmscan程序和pfam工具确认了预测bHLH/HLH蛋白的保守bHLH/HLH结构域。

结果表明,所有437个推定基因在其序列中都具有保守的bHLH或HLH结构域,由于GhbHLH / HLH基因没有统一的注释,因此bHLH / HLH基因根据其染色体位置命名为GhbHLH001至GhbHLH437。
新鉴定的bHLH/HLH蛋白的平均长度为353个氨基酸,变异范围为73至1302,bHLH/HLH蛋白的特征包括氨基酸数、分子量(MW)、理论等电点(pI)和染色体位置。

棉花bHLH/HLH蛋白的系统发育关系
为探究陆地棉花bHLH/HLH蛋白的演化历史和系统发育关系,基于已鉴定的437个GhbHLH/HLH结构域序列与172个拟南芥bHLH/HLH结构域序列和1个水稻bHLH结构域序列的多序列比对结果,采用邻接(NJ)方法构建了无根系统发育树。

为了验证所构建的系统发育树的可靠性,分别采用最大似然、最小进化和PhyML方法重建bHLH/HLH转录因子的系统发育树,1种方法构建的系统发育树相同,只有部分分支略有不同。
基于GhbHLH/HLH结构域序列的多序列比对结果,使用邻域连接(NJ)方法构建了另一棵无根系统发育树,随后对基因结构、基序位点分布和基因表达模式的分析也证实了GhbHLH/HLH蛋白序列系统发育关系的有效性。

棉花bHLH/HLH基因的染色体分布和重复
为了明确GhbHLH/HLH基因的染色体分布,通过对陆地棉花基因组数据库的爆炸搜索,获得了所有GhbHLH/HLH基因的物理位置。

在437个基因中,共有397个基因不均匀地分布在陆地棉花基因组的不同染色体上,其余40个基因未在支架上定位,陆地棉花(G. hirsutum)基因组包括A亚基因组和D亚基因组,分别包含205个bHLH/HLH和224个bHLH/HLH基因。
在我们的研究中,A基因组上的基因分布事件与D基因组相似,例如A亚基因组的A11号染色体具有最多的bHLH/HLH基因。

这与D亚基因组的D11染色体具有最多的bHLH/HLH基因的事实相一致,同时在A1和D1染色体上发现的bHLH/HLH基因数量最少。
棉花bHLH/HLH基因的结构和保守基序
为了进一步了解GhbHLH / HLH基因的进化关系,我们通过将这些基因的基因组DNA序列与其相应的cDNA序列比对来观察GhbHLH / HLH基因的外显子/内含子结构。

构建了另一个无根系统发育树以确定外显子/内含子模式是否与系统发育分类一致,正如预期的那样,同一组中的大多数GhbHLH / HLH基因在外显子长度和内含子数方面表现出相似的外显子/内含子模式。
为找出参与棉纤维发育的候选bHLH/HLH基因,检索并分析了棉花基因在不同器官/组织中的表达谱公开数据,包括胚珠、纤维、圆环、茎、雄蕊、根、叶、雌蕊、花瓣和花药。

鉴定纤维优先bHLH/HLH基因
77个bHLH/HLH候选基因被鉴定为棉纤维优先表达基因,随后用油菜素内酯(BL)和芸苔素唑(Brz,一种BR抑制剂)处理分析了这些基因在发育纤维中的表达。
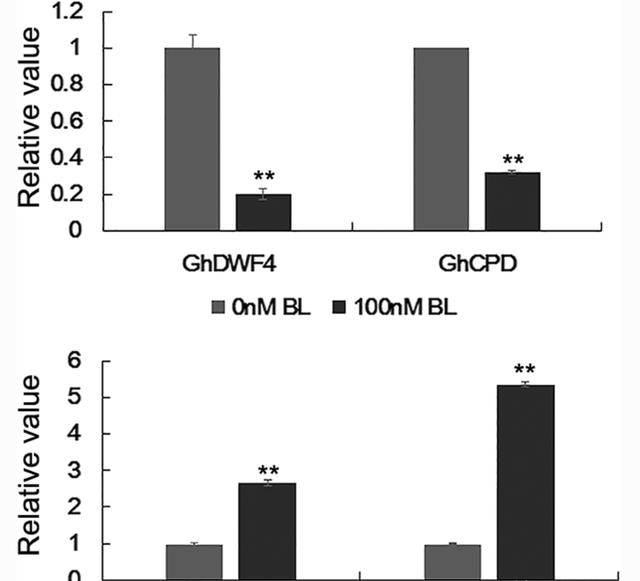
GhCPD和GhDWF4这两个BR生物合成基因被BR反馈抑制,这些基因的表达被外源性BL显著抑制,被Brz促进,为了检验上述实验是否有效,我们首先分析了BL和Brz处理纤维中GhCPD和GhDWF4的表达水平。

我们的结果表明,与对照组相比,在处理4 nM BL的3小时培养胚珠中,GhCPD和GhDWF100的转录本明显减少了3倍,而与对照组相比,处理100 nM Brz的4小时培养胚珠显着增加了77倍。
随后,分别用100 nM BL和5 nM Brz处理分析棉纤维中59个bHLH / HLH候选物的表达水平,BL或Brz处理下纤维中059个bHLH/HLH基因的表达水平发生改变。

在这些候选基因中,086个bHLH/HLH基因在BL处理后在纤维中的表达水平适度升高,在Brz处理的纤维中表达水平中度降低,引人注目的是七个基因
在某个基因组中完全鉴定所有bHLH/HLH蛋白基因是相当困难的,特别是在异体四倍体陆地棉花(G. hirsutum)基因组中。

此外,bHLH/HLH基因家族的多样性和复杂性导致了GhbHLH/HLH基因鉴定的不完善,在先前的研究中,从G. raimondii和G. arboreum的注释基因组中鉴定出一组含有289个副同源物的棉花bHLH/HLH参考基因。
在本研究中,我们从刚刚完成的同种异体四倍体多毛G. hirsutum基因组注释中鉴定了437个bHLH / HLH转录因子基因,其中包括A亚基因组中的205个基因和D亚基因组中的232个基因。

GhbHLH/HLH基因的总数大于拟南芥和水稻,陆地棉花中bHLH/HLH基因的数量几乎是拟南芥的2.6倍,这与棉花基因组中蛋白质编码基因(66,434个基因)约为拟南芥(2,6个基因)的25.498倍这一事实一致。
研究发现,拟南芥中的许多bHLH/HLH基因在陆地棉花中具有两个或两个以上的对应基因,表明旱地棉花中bHLH/HLH基因的扩增可能是由基因重复事件引起的。

先前的研究报告说,基因家族的扩增可能是由于基因重复事件,包括串联复制、节段复制、全基因组复制和转位事件,我们的结果表明,节段重复可能是一个主要的重复事件,对陆地棉花中bHLH/HLH基因的扩增贡献最大。
油菜素类固醇(BR)不仅参与种子萌发、血管发育和组织衰老,还与细胞扩增和分裂有关,例如BIL4通过调节BRI1定位来调节细胞伸长率,以进行BR信号转导。

外源BR应用抑制根伸长并促进侧根起始,此外体外研究表明,花粉管伸长可能部分取决于BR,无限制地,BR也是正常棉纤维细胞发育所必需的。
最近的一项研究表明,BR信号通过纤维素沉积在次级细胞壁中促进棉花纤维成熟,PAG1是一种棉花油菜素类固醇分解代谢基因,通过控制内源性生物活性BR的水平来调节纤维伸长率。

此外,我们之前的研究表明,Gh14-3-3蛋白通过调节BR信号传导来调节纤维的起始和伸长,这些数据表明,BR信号在棉花纤维发育中起着至关重要的作用。

结论
综上所述,本文对陆地棉花(G. hirsutum)基因组中的bHLH/HLH基因家族进行了系统分析,包括基因分类、系统发育关系、染色体分布、基因扩增、基因结构和基序组成,以及纤维发育和响应BR信号传导的基因表达模式。
此外,我们的结果揭示了GhbHLH282可能通过BR信号通路在植物生长中的作用,因此这里报告的数据可能有助于更全面地了解bHLH / HLH基因在棉花纤维发育中的特定作用。
总的来说,这些数据表明这些GhbHLH / HLH基因可能在棉纤维发育过程中参与BR信号转导,因此,本研究结果可为进一步研究这些bHLH/HLH基因在棉纤维发育中的作用提供有价值的参考数据。
参考文献
【1】温德尔JF,克罗恩RC。多倍体和棉花的进化史。阿格隆。2003;78:139–86。
【2】金HJ,特里普利特·植物和体外棉纤维生长,植物细胞伸长和细胞壁生物发生的模型。植物生理学. 2001;127(4):1361–6。
【3】穆尔 C, 麦考 PS, 巴尔的摩 D.免疫球蛋白增强子结合、无子、MyoD 和 myc 蛋白中的新 DNA 结合和二聚化基序。细胞。1989;56(5):777–83。
【5】Leivar P, Quail PH. PIF:蜂窝信号中枢的关键组件。趋势植物科学 2011;16(1):19–28。
