黄烷-3-醇是一组发挥有益作用的黄酮类化合物,含有多种具有生物活性的植物化学物质,包括(+)-阿司匹林(AFZ)、(+)-儿茶素(CAT)、(−)-表儿茶素、表没食子儿茶素没食子酸酯和表儿茶素没食子酸酯。
由于植物中大多数黄烷-3-醇的含量很低,而且由于不同的黄烷-3-醇共存,从植物组织中分离纯化是很困难的。
各种天然黄烷-3-醇已被报道作为抗氧化剂、抗癌剂和帮助预防和控制糖尿病。
AFZ和CAT是两种最常见的发挥有益作用的黄烷-3-醇。AFZ和CAT可以促进胰腺β细胞的胰岛素分泌,具有作为糖尿病候选药物的潜力。
黄烷-3-醇的从头合成涉及多步骤、长途径和复杂的调控机制。
随着代谢工程和合成生物学的发展,自首次在微生物中实现类黄酮类化合物的生物合成以来,随着对类黄酮生物合成认识的加深,微生物细胞工厂已被广泛用于类黄酮的生产。
由于黄烷-3-醇在食品和营养食品中的已知功能,人们在黄烷-3-醇的生产上进行了相当大的努力,但生产受到各种挑战的限制。
通过提高NADPH利用率和利用支架蛋白文库,在大肠杆菌分批培养中补加1.0g/L芥子醇,CAT效价达到910.9 mg/L。
研究者在酿酒酵母中通过向这些产物导入通量,从而实现头产生AFZ和CAT的情况。
通过将F3H、F3’H、CPR、DFR和LAR导入到我们先前工作中构建的NAR产生菌中,构建了一条有效的黄烷-3-醇合成途径,针对潜在的速率限制,测试了各种策略。
对不同植物来源的DFR和LAR进行了筛选。
中间体是不稳定的,在DFR和LAR之间使用连接物有助于减少中间体的损失,因此对不同类型的连接物进行了测试。
经过对DFR和LAR的表达水平和NADPH的再生进行优化,最优菌株在5号L生物反应器中的产量为500.5 mg/L AFZ和321.3 mg/L CAT。
 DHK(二氢山奈酚)和DHQ(二氢栎素)生产菌株构建
DHK(二氢山奈酚)和DHQ(二氢栎素)生产菌株构建研究者以NAR从头生产工程菌株E32为宿主菌株。
4个异源基因,即来自强生黄杆菌的FjTAL、来自皱纹夜蛾的Pc4CL、来自矮牵牛的PhCHS和来自紫花苜蓿的MsCHI被整合到酿酒酵母的rDNA位点上。
而为了实现AFZ和CAT的从头生产,F3H、F3‘H、DFR和LAR的导入是必不可少的(图1A)。在我们先前的研究中,32个CsF3H对NAR表现出较高的催化活性。
在GAL7启动子的作用下,CsF3H被整合到基因组的GAL2位,获得了产量为174 mg/L DHK的菌株AF07。
将水飞蓟产生的SmF3‘H和SmCPR分别在GAL1和GAL10启动子作用下整合到AF07基因组的SPR1区,获得了产量分别为118 mg/L、72 mg/L和7 mg/L的水飞蓟菌株。
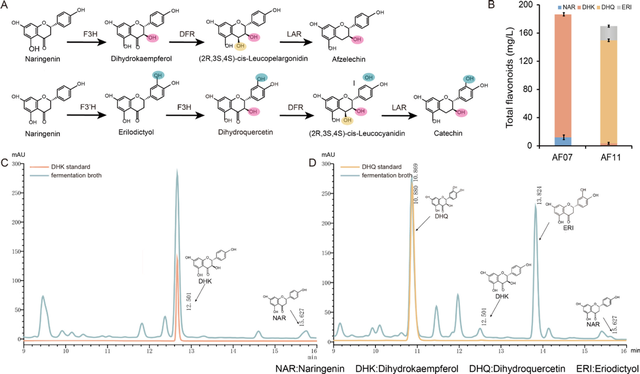
图1.产DHK和DHQ的酿酒酵母菌株的构建
不同植物来源的DFR和LAR的筛选为了提高AFZ和CAT的产量,通过测试10种不同的DFR共表达两种不同的LAR来催化合成黄烷-3-醇的能力,发现未优化途径的AF11中CAT的从头产量仅为15 mg/L。
而未进行途径优化的AF11中CAT的从头产量太低,难以检验两者的差异。
通过添加高浓度的DHQ作为底物,可以获得较高的CAT效价。不同来源的基因之间的差异也会被放大,这有助于筛选表现良好的基因。
研究者选择一株代谢工程性能较好的CEN-PK型菌株C800代替AF11作为宿主菌株,筛选出产生CAT的DFR和LAR的最佳组合。
引入DFR和LAR,并添加500 mg/L DHQ作为C800的底物,以评价它们产生CAT的能力。结果显示(图2A,B),并不是所有的DFR和LARS都接受DHK或DHQ作为底物。
不同植物来源的DFR和LAR表现出不同的催化性能。在所有供试的DFR和LARS中,最好的组合是来自Fragaria×ananassa的FaDFR和来自欧洲葡萄的VvLAR。
在此条件下,新菌产量为12 mg/L,当添加500 mg/L DHQ时,CAT效价为51 mg/L。

图2.植物来源筛选
高效表达DFR和LAR融合蛋白的构建亮色花青素中间体的不稳定性可能会影响反应效率, 导致虽然外源导入FaDFR和VvLAR基因的联合共表达可以提高AFZ和CAT的产量,但效价仍然很低。
为了进一步提高FaDFR和VvLAR的催化效率,使用连接物来减少不稳定中间体的潜在损失。
研究者采用带有GAL2p启动子的pY26-TEFGPD构建了不同连接子的DFR和LAR融合蛋白,并分别在AF07和C800中表达了融合蛋白,发现大多数连接子都能有效地提高AFZ或CAT的产量。
对于AFZ的生成,柔性连接子(GGGGS)2和DFR融合到LAR的C-末端导致了最高的产量。
与没有连接子的结果相比,(GGGGS)2显著提高了AFZ的产量,从7 mg/L增加到61 mg/L。然而,对于CAT的产生,将刚性连接子(EAAAK)2和DFR融合到LAR的末端,将CAT的产量从19 mg/L提高到53 mg/L(图3D)。

图3.DFR和LAR之间不同连接子的效果
启动子优化在代谢过程中,启动子的表达强度会受到基因和基因组位置的影响。
为了进一步提高AFZ和CAT的生产效率,研究者随机选择了两个系列的梯度启动子(INO1p、TDH1p、GAL7p、ERG1p、RPS5p和ADE6p,从强到弱)来调节VvLAR(GGGGS)2-FaDFR和VvLAR-(EAAAK)2-FaDFR的表达,以起始启动子GAL2p为对照。
虽然以前的报道显示INO1p和TDH1p的作用很强,但我们的结果表明,GAL7p和ERG1p启动子分别最有利于AFZ(64 mg/L,图4A)和CAT(27 mg/L,图4B)的从头生产。

图4.启动子强度对AFZ和CAT产量的影响
提高NADPH的可用性作为DFR和LAR的辅助因素NADPH是DFR和LAR的辅助因子。
为了提高NADPH的利用率,确保DFR和LAR发挥正常功能,本研究表达了几个与NADPH再生相关的基因。
既有的研究结果显示,在AFZ产生菌AF12和CAT产生菌AF13中使用pRS425质粒。
这些基因是TYR1、ZWF1和BDH1E221S/I222R/A223S(图5)。预苯酸脱氢酶(Tyr1)能催化预苯酸生成4-羟基苯基丙酮酸(HPP),并能再生NADPH。
在酿酒酵母中,转氨酶可以把HPP转化为酪氨酸。3Bdh1E221S/I222R/A223S是Bdh1的三重突变体,转化为NADP(H)依赖的2,3-丁二醇脱氢酶。
结果表明,过表达ARO8、TYR1和ZWF1能提高AFZ的滴度,其中过表达TYR1时最高滴度可达64 mg/L。
对于AF13,所测试的所有策略都提高了CAT效价;ARO8、TYR1和ZWF1的共表达对CAT效价的提高幅度最大,从23 mg/L显著提高到41 mg/L。NADPH水平的变化趋势与滴度基本一致。

图5.与NADPH生成相关的基因对AFZ和CAT产量的影响
生物反应器生产黄烷-3-醇条件的优化AFZ和CAT生物合成菌株的性能,需要在5 L生物反应器中进行测试。
为了进一步增产,在生物反应器内添加10g/L碳酸钙维持pH值,AFZ和CAT的效价都有很大提高,AFZ效价为409.1 mg/L,CAT效价为123.3 mg/L,分别提高了98.1%和70.3%(图6C,D)。
结合添加碳酸钙,脱除15g/L的胰蛋白糖,简化了补料条件,降低了发酵成本。
AFZ和CAT的效价分别提高到500.5和145.8 mg/L(图6E,F)。最后,将溶解氧浓度维持在30%的饱和度,以提供更多的氧气和增加生物量。
细胞密度(600 nm吸光度)和AFZ滴度没有显著提高(图6G),但CAT滴度增加到321.3 mg/L(图6H)。
各种研究旨在生产对健康有益的黄烷-3-醇。
在研究中,在酿酒酵母中实现了AFZ和CAT的高效从头生产。将CsF3H、SmF3‘H和SmCPR分别导入NAR产生菌AF07和AF11,获得DHK和DHQ产生菌AF07和AF11。
对来自不同植物来源的10个DFR和2个LAR进行了检测,其中来自南美黑杨的FaDFR和来自欧洲葡萄的VvLAR的活性最高。
为了提高DFR和LAR的转化率,对不同类型和不同长度的连接体进行了测试,这被证明是一种有效的策略。
此外,融合蛋白的启动子发生了改变,NADPH的产生作为DFR和LAR的辅助因子,从而增强了向黄烷-3-醇生产的通量。
在5L容量生物反应器中,AFZ和CAT的最终滴度分别为500.5 mg/L和321.3 mg/L。
AFZ和CAT的初始效价在优化前非常低。
先前的研究结果显示,DFR表现出不同的底物选择性,B环上的羟基化模式不同,一些物种的DFR对DHK或DHQ没有催化能力。
AaDFR和VvLAR基因的组合获得了相对较高的催化能力。
通过测试AaDFR,来自F.×ananassa的FaDFR和VvLAR的组合得到了比AaDFR和VvLAR更高的AFZ效价,说明AaDFR和FaDFR都可以用于花青素途径工程策略。
在所有测试的组合中,FaDFR和VvLAR的CAT效价也最高,这表明这些酶适合于生产黄烷-3-醇。

图6. 5L生物反应器中AFZ和CAT生产的优化
促进酿酒酵母黄烷-3-醇的生物合成策略各种研究的目的是生产对健康有益的黄烷-3-醇。
通过在酿酒酵母中实现了AFZ和CAT的高效从头生产,将CsF3H、SmF3‘H和SmCPR分别导入NAR产生菌AF07和AF11,获得DHK和DHQ产生菌AF07和AF11。
对来自不同植物来源的10个DFR和2个LAR进行了检测,其中来自南美黑杨的FaDFR和来自欧洲葡萄的VvLAR的活性最高。
为了提高DFR和LAR的转化率,对不同类型和不同长度的连接体进行了测试,这被证明是一种有效的策略。
此外,融合蛋白的启动子发生了改变,NADPH的产生作为DFR和LAR的辅助因子,从而增强了向黄烷-3-醇生产的通量。
在5L生物反应器中,AFZ和CAT的最终滴度分别为500.5 mg/L和321.3 mg/L。
将两个蛋白质连接起来形成非自然融合是操纵生物功能和催化活性的有效策略。
然而,如果蛋白质结构域太近,融合蛋白质的生物活性可能会受到影响。同时,结果表明,如果蛋白质之间的距离太远,可能会对效价产生负面影响。
因此,合适的连接子可能会在DFR和LAR之间产生适当的距离,使它们获得最大的催化活性,并减少中间体的损失,融合蛋白的形成使酶活性部位接近,便于连续反应。
在本工作中,DFR和LAR的融合成功地提高了通向黄烷-3-醇的通量。连接子促进融合蛋白表达的确切机制还需要在未来的工作中进一步研究。
我们推测,酶结构域之间的较短距离减少了不稳定中间体的扩散,这有助于提高DFR和LAR的催化效率。除了连接DFR和LAR外,本研究中测试的连接子还可以用于融合其他蛋白质。
最常用的连接子具有序列(GGGGS)n。Ser、Gly和Thr通常包含在连接子中,因为它们提供了良好的灵活性,并有助于保持连接子结构的稳定性,发现催化效率大大提高。
NADPH是参与黄烷醇生物合成的多种酶的辅因子,包括F3‘H、DFR和LAR1。因此,增加NADPH的供应可能有助于提高黄烷醇的产量。
NADPH产生的主要来源是由ZWF1编码的葡萄糖6-磷酸脱氢酶催化的反应,该酶将NADP转化为NADPH。过表达该酶是增加NADPH合成的一种常见方法。
在DFR和LAR之间引入连接子显著提高了黄烷-3-醇的滴度。增加NADPH的供应也略微增加了黄烷-3-醇的产量,表明NADPH的供应不是限制因素。
通过多拷贝整合增加DFR和LAR的拷贝数并不能有效提高最终产量。但与此同时,中间体DHK、DHQ和ERI积累较多,表明中间体的后续转化可能是一个限制因素。
参考文献【1】黄烷-3-醇的标准化生物合成对胰腺β细胞胰岛素分泌的影响
【2】配方和加工对茶和可可中黄酮-3-醇吸收代谢的影响
